



User Protocol 01:
Determination of the Chemotactic Behavior of Campylobacter jejuni by using μ-Slide Chemotaxis
Elgamoudi, Bassam and Ketley, Julian
Department of Genetics, University of Leicester, University Road, Leicester LE1 7RH, UK
*For correspondence: ket@le.ac.uk
Campylobacter jejuni is an important gastrointestinal pathogen of humans. While pathogenicity of C. jejuni remains poorly understood, chemotactic motility has been shown to be involved in cell invasion and colonisation of the gastrointestinal tract. The measurement of migration by chemotaxis assays can be complicated because metabolic consumption of chemoeffector may create a secondary gradient that the bacteria can sense. Tests like the hard agar plug (HAP) and capillary assays are standard methods used to measure bacterial chemotactic responses toward attractants and repellents. However, in C. jejuni these assays may lead to erroneous results, such as excessive experimental variation, unsuitability for studying chemorepellents, and false positive responses. The aim of this study was to establish a reproducible method to measure chemotactic responses of C. jejuni quantitatively over a short time-period.
Here we describe the use of the μ-Slide Chemotaxis chamber (ibidi GmbH, Gräfelfing, Germany) to measure the chemotactic responses of C. jejuni. The μ-Slide Chemotaxis microscopic slide has two reservoirs, one contains bacteria and the other contains chemoattractants, and migration in response to the formed chemoattractant gradient is monitored by microscopy and viable cell counts. The application of the μ-Slide Chemotaxis chamber combined with a C. jejuni-adapted chemotaxis assay demonstrated that C. jejuni has preferential patterns of chemotaxis, where serine is preferred to proline, then glutamate and lastly aspartate. The modified μ-Slide Chemotaxis chamber allowed detection of the chemotactic responses, as well as tracking individual cells to study motility patterns.
1. Cell Culture Media, Buffer, and Solutions
- Strains used in this study
C. jejuni NCTC11168 (National Collection of Type Culture, Colindale, London, UK), C. jejuni 81116 flaAB (non-motile) and C. jejuni NCTC11168 cheY (non-chemotactic) (Kanungpean et al. 2011) were used in the study. C. jejuni cells were grown under microaerobic conditions (85% N2, 10% CO2, and 5% O2) at 42ºC. Mueller-Hinton Agar (MHA) and Mueller-Hinton Broth (MHB) supplemented with trimethoprim and vancomycin, which specifically selects for C. jejuni were used. Bacterial cells were sub-cultured by swabbing onto fresh MHA plates for overnight in the VAIN at 42ºC under the microaerophilic environment (85% N2, 10% CO2 and 5% O2.). After that, colonies were visible and ready to be used in experiments. - MH broth and Agar
Sterile MH broth was made using 8.4 g MH broth powder (Oxoid, Basingstoke, UK; 0.15% starch, 0.6% meat infusion and 1.75% casein hydrolysate) dissolved in 400 ml distilled water and autoclaved at 121°C for 15 minutes. MH Agar was made using 15.2 g MH agar / 400 ml distilled water (Oxoid, Basingstoke, UK) and sterilized by autoclaving. - Buffered Saline (BS) medium
BS-agar medium was prepared by dissolving 0.1, 0.8 or 4% (w/v) Bioagar (Biogene Ltd, UK) per 100 ml of BS. BS solution was prepared by dissolving one tablet of BS (ICN Biomedicals) in 100 ml of dH2O. The medium was sterilized by autoclaving. - L-amino acids
Serine, aspartate (potassium), glutamate (monosodium) and proline. All were obtained from Sigma Aldrich. - Low melting temperature (LMT) agarose (Sigma)
1% of LMT agarose was prepared in BS (1 x BS with 1% (w/v) agarose). - Trimethoprim (Final concentration (5 µg ml-1) and Vancomycin (Final concentration (10 µg ml-1) (TV)
All stock solutions were stored at 4°C in the dark until used. All antibiotics were purchased from Sigma-Aldrich and filter sterilized using a 0.2 μm Acrodisc prior to use (PALL Life Sciences, Portsmouth UK).
2. Equipment
- Olympus CellR/ScanR microscope system
- Vortex (Vortex-Genie 2)
- Water bath at 65°C
- Eppendorf centrifuge 5810R
- Spectrophotometer
- µ-Slide Chemotaxis, 80326, ibidi GmbH, Gräfelfing, Germany
- Variable-atmosphere incubator (VAIN)
3. Procedure
µ-Slide Chemotaxis, consisting of two separate opposing liquid chambers or reservoirs, divided by a 1-mm narrow liquid transition zone was used for this assay (Figure 1). A 1% LMT agarose gel was applied to the transition zone. The first reservoir was completely filled with the bacterial cell suspension, the second reservoir with chemoeffector. Three biological repeats of all experiments, including controls, were performed. The numbers of bacteria which transmigrated from the first reservoir into the transition chamber or the second reservoir, with/without chemoeffectors, were observed by using cell counts.
- Incubate the slides in the VAIN under microaerobic conditions (5% O2, 10% CO2 and 85% N2) for 30 minutes to allow gas exchange.
- Fill the transition zone with 10 µl of 1% of LMTagarose gel and leave at room temperature for 10 minutes.
- Dilute chemoattractants in 1% of LMTagarose gel (1:3 vol:vol). Apply 60 µl into one reservoir and leave at room temperature for 10 minutes. (See Note 2)
- Harvest C. jejuni cells in late stationary phase from MHA plates using 2 ml of BS. Centrifuge at room temperature at 6000×g for 5 minutes or full speed for one minute.
- Wash cells with BS and centrifuge again.
- Resuspend the pellet in 2 ml of BS and adjust the OD600 to 1.
- Mix 500 µl of cells with 1% LMTagarose gel. Adjust the cell concentration to a final OD600 of 0.5. See Note 3.
- Load 60 µl of the diluted cell culture into the second reservoir.
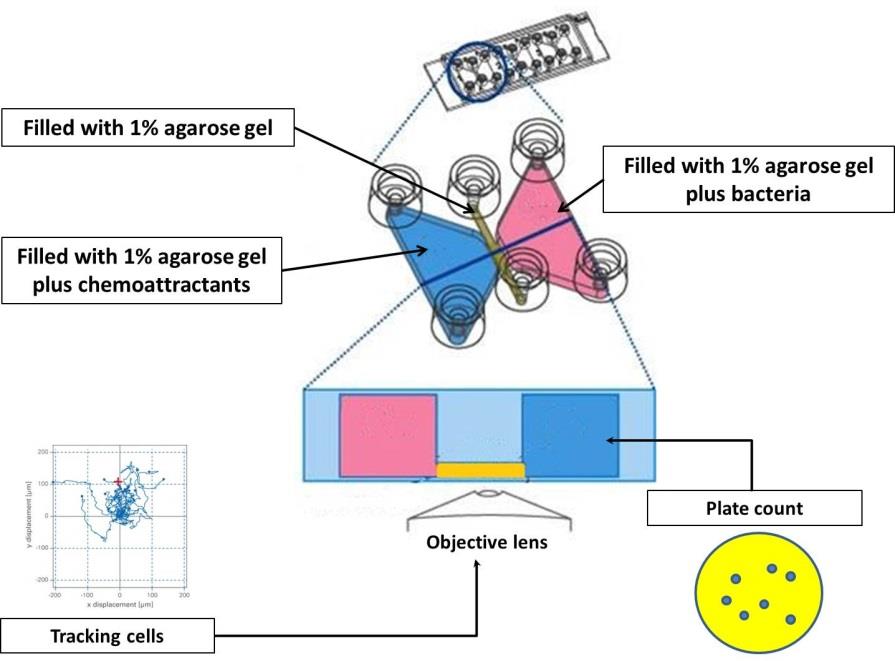
Figure 1. An outline of the chemotaxis slides assay. Briefly, two large reservoirs (volume 60 µl in each) are connected by a narrow observation area (transition zone that is filled with 1% agarose gel). The chemoeffectors-agarose gel is loaded into the first reservoir and then finally, the C. jejuni cells embedded in agarose gel are inserted into the second reservoir. After 3 hours of incubation time, contents of chemoeffector reservoir were serially diluted and spotted out onto MHA plates. The plates were incubated for 48 hours after which viable counts were performed (Adapted from ibidi GmbH).
3.1 Analysis using viable counts (Colony Forming Units)
- Incubate for 3 hours in the VAIN at 42°C under microaerobic conditions.
- After that, a 20 µl sample was taken from the chemoeffectors reservoir.
- Dilute the 20 µl samples serially in MHB to the following dilutions: 10-1, 10-2, 10-3, 10-4, 10-5 and 10-6.
- Plate a 10 µl drop of the dilutions onto MHA plates to obtain viable counts.
- Incubate in the VAIN for two days at 42°C.
- Determine the number of colonies forming units (CFU/ml).
3.2 Representative Results from Viable Counts (Colony Forming Units)
The behavior of wild-type NCTC 11168 towards 0.5 M of proline, serine and aspartate were carried out. The statistical analysis shows that cells have a significantly higher chemotactic response to serine (6 X 107 CFU/ml, p= 0.0069), proline (4.1 X 107 CFU/ml, p< 0.0001) and aspartate (7 X 106 CFU/ml, p= 0.0017) compared to the BS control (8.2 X 105 CFU/ml).
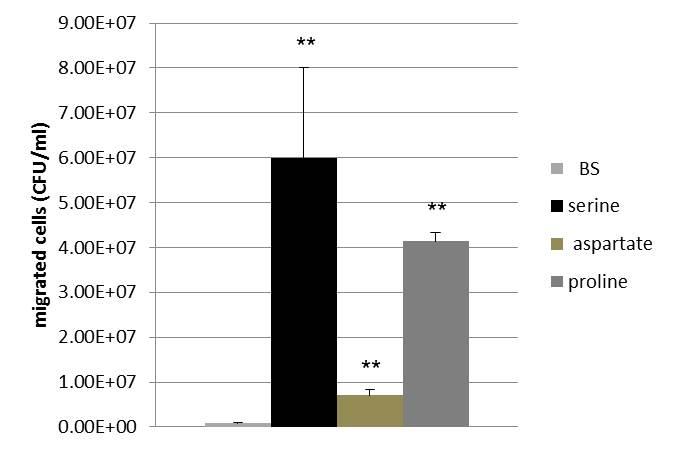
Figure 2. Chemotactic behavior of wild-type NCTC 11168 towards 0.5 M of proline, serine and aspartate. Data presented as Mean ±S.D. ** significant differences.
3.3 Analysis with Cell Counts by Microscopy
- Incubate for 3 hours in the VAIN at 42°C under microaerobic conditions.
- Take photographs in the chemoeffector reservoir with an objective lens 60x or 100x.
- Examine ten fields of view in the chemoeffector reservoir.
- Count transmigrated cells from the photographs using the ImageJ plugin MtrackJ.
3.4 Representative Results with Cell Counts by Microscopy
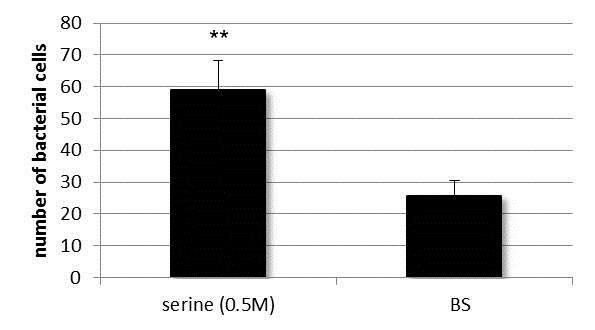
Figure 3. The number of transmigrated cells of wild-type C. jejuni after 3 hours of incubation in microaerobic condition at 42°C. Cells have significantly transmigrated into the serine-containing reservoir, while control experiments with BS permitted unhindered passage of bacteria to the second reservoir. Data presented as Mean ±S.D. ** significant differences.
3.5 Analysis with Single Bacteria Tracking
- Incubate for 1 hour in the VAIN at 42°C under microaerobic conditions.
- Acquire bright field images with a 60× or 100× objective lens and a frame rate of 120 frames/s for 5 s.
- Examine ten fields of view in the transition zone.
- Analyze the motility pattern of at least ten bacteria per strain in each experiment.
- The ImageJ software plugins MtrackJ (http://www.imagescience.org/meijering/software/mtrackj/) and Chemotaxis and Migration Tool (ibidi GmbH, Martinsried, Germany) are used to track the single-bacterium cells.
- Verify the data by performing a student’s t-test.
3.6 Representative Results with Single Bacteria Tracking
The motility pattern of wild-type C. jejuni was a smooth swimming zigzag-like movement toward 0.5 M of serine which represents frequent changes of swimming direction being observed. As demonstrated in Figure 4, the ∆cheY mutant had lost the typical swimming pattern of the wild type and swam in a rather straight manner in almost one direction as was seen in the tracks of non-chemotactic mutant (Marchant 1999, Takata et al. 1992).
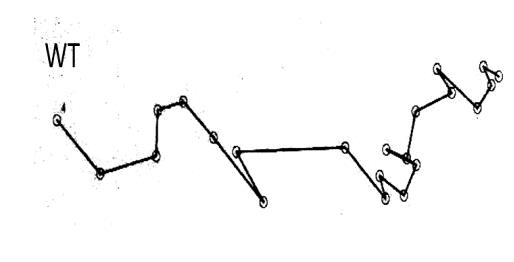

Figure 4. Representative motility pattern of wild-type C. jejuni (WT) and ∆cheY. The figure shows the results of the tracking analysis of individual cells of wild type and ∆cheYmutant response to 0.5 M of serine for 5s.
4. Conclusion
The protocol described in this paper shows a promising reproducible result in short time-period. The results demonstrated that C. jejuni has preferential chemotactic patterns, where serine preferred to proline, glutamate, and lastly aspartate. Moreover, motility patterns indicate that mutant cheY cells swim in a straight manner and cannot change the direction as described before by Yao et al. (1997). In contrast, wild-type shows the typical straight movement with more changing directions.
5. Acknowledgements
This work supported by the Ministry of Higher Education and Scientific Research, Libya. We acknowledge Dr. Kees Straatman from the University of Leicester for his help with microscopy preparation and Professor Victoria Korlik and Elias Horn for their helpful suggestions.
6. Notes
- This protocol is a part of a Ph.D. thesis (Elgamoudi 2016).
- Take care not to introduce any air bubbles into the whole chamber system.
- Optionally, add 0.1% Tween-20 to avoid cell adherence to the slide's surfaces as recommended (Armitano et al. 2011, Baraquet et al. 2009).
- Non-motile (flaA-) and non-chemotactic (cheY-) mutants should be used as controls.
7. References
Armitano, J., Baraquet, C., Michotey, V., Mejean, V. and Jourlin-Castelli, C. (2011) The chemical-in-muwell: a high-throughput technique for identifying solutes eliciting a chemotactic response in motile bacteria. Res Microbiol,162(9), pp. 934-8.
Baraquet, C., Théraulaz, L., Iobbi Nivol, C., Méjean, V. and Jourlin Castelli, C. (2009) Unexpected chemoreceptors mediate energy taxis towards electron acceptors in Shewanella oneidensis. Mol Microbiol,73(2), pp. 278 -290.
Elgamoudi, B. A. (2016) The role of transducer-like proteins in Campylobacter jejuni. Unpublished Ph.D thesis, University of Leicester.
Kanungpean, D., Kakuda, T. and Takai, S. (2011) False positive responses of Campylobacter jejuni when using the chemical-in-plug chemotaxis assay. The Journal of veterinary medical science / the Japanese Society of Veterinary Science,73(3), pp. 389 -391.
Marchant, J. (1999) Studies into the chemotaxis of Campylobacter jejuni. Unpublished PhD, Leicester, Leicester.
Takata, T., Fujimoto, S. and Amako, K. (1992) Isolation of nonchemotactic mutants of Campylobacter jejuni and their colonization of the mouse intestinal tract. Infect Immun,60(9), pp. 3596 -3600.
Yao, R., Burr, D. H. and Guerry, P. (1997) CheY mediated modulation of Campylobacter jejuni virulence. Mol Microbiol,23(5), pp. 1021 -1031.
This User Protocol is an ibidi peer-reviewed protocol from an actual user. ibidi does not guarantee its functionality or reproducibility. For this User Protocol, ibidi provides only limited support. Please contact the corresponding author for detailed information.
For research use only.